| Accession: | |
|---|---|
| Functional site class: | SH3 domain ligands |
| Functional site description: | The SH3 domain is one of the best characterized protein domains. SH3 domains are involved in a wide-range of important cellular processes including intracellular signaling, cytoskeletal rearrangements and cell movement, cell growth and immune responses. They bind to proline-rich sequences with moderate selectivity. Early studies identified “PxxP” as a core conserved sequence motif for SH3 binding. These motifs are referred to as canonical binders, among which class I and class II ligands are distinguished based on their orientation. Since then, SH3 domains recognizing partners with multiple atypical SH3 binding motifs have also been described. |
| ELMs with same func. site: | LIG_SH3_1 LIG_SH3_2 LIG_SH3_3 LIG_SH3_4 LIG_SH3_PxRPPK_7 LIG_SH3_PxxDY_5 LIG_SH3_PxxPPRxxK_8 LIG_SH3_PxxxRxxKP_6 |
| ELM Description: | this is the motif recognized by class II SH3 domains |
| Pattern: | P..P.[KR] |
| Pattern Probability: | 0.0011112 |
| Present in taxons: | Bos taurus Gallus gallus Homo sapiens Komagataella pastoris Mus musculus Opisthokonta Rattus norvegicus Saccharomyces cerevisiae |
| Interaction Domains: |
|
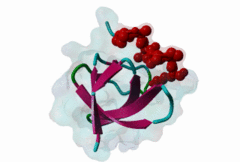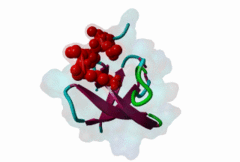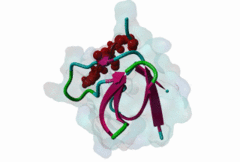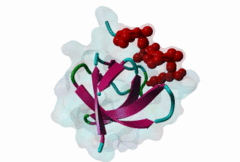
SH3 (SRC Homology 3) domains constitute one of the largest protein domain families with over 300 representatives in the human proteome and 30 in the yeast proteome. SH3 domains are protein recognition modules that typically function in the assembly of signalosomes and signal transduction (Zarrinpar,2003) in signaling pathways, such as cell growth regulation, endocytosis and remodeling of the cytoskeleton. They are small protein interaction modules consisting of only 60 amino acids. At the secondary structure level, the domain displays a beta-sandwich arrangement of five beta-sheets, 3 loops and a short 310 helix (Saksela,2012). The classical SH3 ligand binding site is made up by two hydrophobic pockets and a negatively charged one usually called the specificity pocket, formed by the RT and the n-Src loops. SH3 domains generally recognise proline-rich motifs forming poly-proline type helices (PPII helixes) when bound to the SH3 (Aitio,2008). Most of the known and well-studied ligands of SH3 domains have the “PxxP” core motif. The ligands that contain the “PxxP” minimal sequence are now referred to as canonical or typical SH3 binding motifs. Based on the extensive searches for SH3 binding motifs, a classification system was established where Class I (LIG_SH3_1) and Class II (LIG_SH3_2) ligands were distinguished (Fernandez-Ballester,2004). Because of the pseudo-symmetrical nature of the PPII helix, the PXXP-binding site can recognize peptides in both orientations by using two different binding modes. In both cases the prolines make contact with the two hydrophobic pockets, but the orientation of the peptide will be determined by the position of the charged residue binding to the specificity pocket; the motif description is “(R/K)xxPxxP” for class I and “PxxPx(R/K)” for class II (Aitio,2008). A key conserved surface Trp residue in the SH3 binding pocket is known to adopt two different orientations that, in turn, determine the type of ligand (I or II) specifically recognized by the domain. Interestingly, some of the SH3 domains are capable of binding ligands both in Class I and Class II orientation (Fernandez-Ballester,2004). Interestingly, motif binding by at least a subset of SH3 domains seems to be regulated by receptor tyrosine kinases (RTKs) through phosphorylation of a conserved C-terminal Tyr residue within the domain that disturbs motif binding and thus leads to the collapse of the associated signaling networks (Dionne,2018). The canonical Class I and II motifs are recognized by diverse SH3 domains. However, a large-scale study on human SH3 domains showed that almost half of the investigated domains exhibit atypical binding specificities with no “PxxP” core (Teyra,2017). The hitherto identified atypical SH3 motifs include the “PxxDY” (LIG_SH3_PxxDY_5) (Li,2005; Kaneko,2008; Saksela,2012), different “RxxK-type” motifs (LIG_SH3_PxxxRxxKP_6; LIG_SH3_PxRPPK_7; LIG_SH3_PxxPPRxxK_8) (Liu,2003; Lewitzky,2001; Harkiolaki,2009; Lewitzky,2004), Px[PA]xPR (LIG_SH3_CIN85_PxpxPR_1) (Kurakin,2003; Rouka,2015), and “RKxxYxxY” (Kang,2000). These atypical recognition specificities are usually associated with only a specific subset of the SH3-containing proteins, which often belong to specific processes/pathways. “PxxDY'' motif binding has only been identified for a relatively few SH3 domain-containing proteins, including Nck1 (P16333) and Eps8L1 (Q8TE68). Nck1 is an adaptor protein functioning in signal transduction between diverse membrane receptors and the cytoskeleton (Li,2001). During the activation of T-cell receptors (TCRs), Nck1 binds to the “PxxDY'' motif of the CD3ε subunit (32317279). In a phage display screening of 296 human SH3 domains only the Nck1, Nck2, Eps8, Eps8L1, Eps8L3 and Eps8L2 proteins were identified as binders of the motif in Cd3ε, indicating that it is highly specific (Kesti,2007). Besides CD3ε, e3b1/abi-1 and US6NL were also demonstrated to have a functional “PxxDY” motif that binds to the SH3 of Eps8 (Mongiovi,1999). Interestingly, two E.coli effector proteins, namely NleH1 and NleH2, have been identified as Eps8 binders. During infection, probably due to being bound by NleH1 and NleH2, Eps8 shows an altered localization pattern within the cytoplasm that might compromise the formation of new microvilli (Pollock,2022). RxxK-type SH3-binding motifs are specifically recognized by the second (C-terminal) SH3 domains of GRB2 (P62993) and GRAP2 (also called GADS (O75791)) (Liu,2003; Lewitzky,2001; Harkiolaki,2009; Lewitzky,2004), and the two STAM proteins, STAM1 (Q92783) and STAM2 (O75886) have also been described to bind certain RxxK motifs (Kato,2000). RxxK motif-mediated interactions are typically involved in the signaling of T-cell and B-cell receptors, as well as receptor tyrosine kinases. The RxxK motifs are highly versatile: there is a canonical version “PxxxRxxKP” (LIG_SH3_PxxxRxxKP_6) and some variants where the RxxK is preceded by a PxxP motif in an overlapping “PxRPxK” (LIG_SH3_PxRPPK_7)(Harkiolaki,2009) or non-overlapping “PxxPxRxxK” arrangement (LIG_SH3_PxxPPRxxK_8; Lewitzky,2004). There is also a variant described where the R and K residues are placed further apart “RxxxxK” (Harkiolaki,2009). There is at least one available structure for all these variants, therefore the differences in secondary structures adopted in the bound peptides as well as in the contacts established with the SH3 pockets/residues are well-understood (Harkiolaki,2009). Most known RxxK motif-containing proteins, such as LCP2/SLP-76, GAB1, 2 and 3, B-cell linker protein (BLNK), STAM-binding protein and Ubiquitin carboxyl-terminal hydrolase 8 (mUBPY) employ the canonical RxxK for SH3 binding. Some proteins, such as GABs, employ more than one RxxK type, while, for instance, MAP4K1/HPK1 employs the combined motif “PxxPxRxxK” for binding to the C-terminal SH3 of GADS (Lewitzky,2004). |
(click table headers for sorting; Notes column: =Number of Switches, =Number of Interactions)
Please cite:
ELM-the Eukaryotic Linear Motif resource-2024 update.
(PMID:37962385)
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement