DOC_PP1_MyPhoNE_1
| Accession: | |
|---|---|
| Functional site class: | PP1-docking motif MyPhoNE |
| Functional site description: | Protein phosphatase-1 (PP1), an enzyme that catalyzes dephosphorylation of proteins, is ubiquitously expressed and highly conserved in eukaryotes. It plays a regulatory role in a wide range of cellular processes, including gene transcription, protein synthesis, cell cycle progression, muscle contraction, and neuronal signalling. The PP1 apoenzyme is a single catalytic domain that can interact with more than 200 regulators, converting it into hundreds of highly specific holoenzymes. The catalytic site of PP1 is at the intersection of three potential docking motif-binding regions: the acidic, hydrophobic and C-terminal grooves (Peti,2013). Most regulatory proteins interact with PP1 at the catalytic site via the RVXF docking motif (DOC_PP1_RVXF_1) but docking motifs such as the SILK motif (DOC_PP1_SILK_1) and the MyPhoNE motif (DOC_PP1_MyPhoNE_1) also play essential roles in regulating PP1 activity and substrate specificity (Hendrickx,2009). |
| ELMs with same tags: |
|
| ELM Description: | The MyPhoNE (Myosine Phosphatase N-terminal Element) motif, generally found N-terminal to an RVxF motif, mediates docking of regulatory proteins to the catalytic subunit of PP1 (PP1c). The peptide is defined by eight amino acid residues and adopts a five-turn alpha helix that interacts with a hydrophobic cleft on the surface of PP1c (1S70) (Terrak,2004). The first position of the motif is invariantly occupied by arginine. The second position is not defined as this residue points away from the binding site, however proline is likely not allowed in this position as this would disrupt the helical conformation. The third position either contains a glutamic acid, a glutamine or an aspartic acid residue. Conservation in this position might be due to an intra-peptide interaction with the side chain of the residue in position 6 or 7, of which at least one always contains a lysine or arginine residue. Such an interaction might stabilize the helical conformation. The fourth position is invariantly occupied by a glutamine, which makes important hydrogen bonds, while a specific hydrophobic residue, either valine, leucine, or isoleucine, is always found in the next position. For position 5, a hydrophic amino acid is needed (valine, isoleucine or leucine). Finally, the last position requires either a tyrosine or tryptophan residue. |
| Pattern: | R[^P][DEQ]Q[VIL]([RK][^P]|[^P][RK])[YW] |
| Pattern Probability: | 4.405e-07 |
| Present in taxon: | Metazoa |
| Interaction Domain: |
Metallophos (PF00149)
Calcineurin-like phosphoesterase
(Stochiometry: 1 : 1)
|
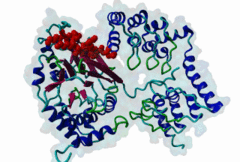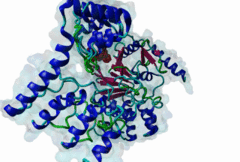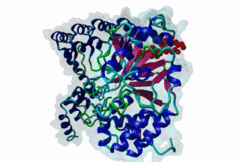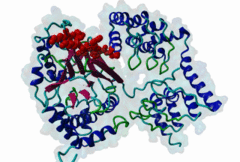
Protein Ser/Thr phosphatases 1-7 (PP1-7) belong to the superfamily of phosphoprotein phosphatases (PPPs) and catalyze protein dephosphorylation in eukaryotes by hydrolyzing Ser/Thr-linked phosphate ester bonds (Heroes,2013). A major Ser/Thr protein phosphatase in eukaryotic cells is Protein phosphatase-1 (PP1). While the yeast Saccharomyces cerevisiae has one PP1 gene (glc7), Mammalia contain four PP1 isofroms encoded by three highly related genes (alpha, beta/delta and gamma), and alternative splicing generating the gamma1 and gamma2 isoforms (Moorhead,2007). Functional PP1 is needed in context of regulating cell cycle progression, protein synthesis, muscle contraction, gene transcription, neuronal signaling, and carbohydrate metabolism (Peti,2013). The activity of PP1 is regulated by a large number of PP1-interacting proteins (PIPs), which can be substrates, inhibitors or targeting subunits of PP1 (Bollen,2010). Binding of these PIPs to PP1 is mediated by different docking sites for docking motifs present in the PIPs. As part of the catalytic subunit of PP1 a hydrophobic, acidic and C-terminal groove have been identified. Over 100 mammalian PP1 interactors have been found, with short and degenerate PP1 docking motifs. Most of the PIPs interact with PP1 via an RVxF motif (DOC_PP1_RVXF_1). This five-residue motif binds as an extended beta-strand 20 Angstroms away from the active site to a hydrophobic groove. In addition to the RVxF motif, some PIPs contain a SILK motif (DOC_PP1_SILK_1). This four-residue motif is always N-terminal to the RVxF motif and essential for the function of Inhibitor-2 (Q9V3C7). It acts as an anchoring motif without changing the conformation of PP1 (Hendrickx,2009). Another N-terminal binding element is the myosin phosphatase N-terminal element or MyPhoNE motif. This motif is present in the myosin phosphatase targeting subunit MYPT1 (O14974) and adopts a five-turn alpha helix that binds to a hydrophobic cleft on the surface of PP1. Similar to the SILK motif, it is generally located N-terminal to the RVxF motif (Bollen,2010). |
-
Structural basis of protein phosphatase 1 regulation.
Terrak M, Kerff F, Langsetmo K, Tao T, Dominguez R
Nature 2004 Jun 17; 429 (6993), 780-4
PMID: 15164081
-
Emerging roles of nuclear protein phosphatases.
Moorhead GB, Trinkle-Mulcahy L, Ulke-Lemee A
Nat Rev Mol Cell Biol 2007 Mar; 8 (3), 234-44
PMID: 17318227
-
Docking motif-guided mapping of the interactome of protein phosphatase-1.
Hendrickx A, Beullens M, Ceulemans H, Den Abt T, Van Eynde A, Nicolaescu E, Lesage B, Bollen M
Chem Biol 2009 Apr 24; 16 (4), 365-71
PMID: 19389623
-
The extended PP1 toolkit: designed to create specificity.
Bollen M, Peti W, Ragusa MJ, Beullens M
Trends Biochem Sci 2010 Aug 06; 35 (8), 450-8
PMID: 20399103
-
Structural basis for protein phosphatase 1 regulation and specificity.
Peti W, Nairn AC, Page R
FEBS J 2013 Jan 29; 280 (2), 596-611
PMID: 22284538
-
The PP1 binding code: a molecular-lego strategy that governs specificity.
Heroes E, Lesage B, Gornemann J, Beullens M, Van Meervelt L, Bollen M
FEBS J 2013 Jan 29; 280 (2), 584-95
PMID: 22360570
(click table headers for sorting; Notes column: =Number of Switches, =Number of Interactions)
| Acc., Gene-, Name | Start | End | Subsequence | Logic | #Ev. | Organism | Notes |
|---|---|---|---|---|---|---|---|
| Q9BZL4 PPP1R12C PP12C_HUMAN |
22 | 29 | AAARERRREQLRQWGARAGA | TP | 1 | Homo sapiens (Human) | |
| Q90623 PPP1R12A MYPT1_CHICK |
10 | 17 | ADAKQKRNEQLKRWIGSETD | TP | 1 | Gallus gallus (Chicken) | |
| Q5SQS7 SH2D4B SH24B_HUMAN |
32 | 39 | ILFYKMREEQLRRWKERETW | TP | 1 | Homo sapiens (Human) | |
| Q9H788 SH2D4A SH24A_HUMAN |
32 | 39 | ILFFKMREEQIRRWKEREAA | TP | 1 | Homo sapiens (Human) | |
| Q96T49 PPP1R16B PP16B_HUMAN |
30 | 37 | RAAQKRRAQQLKKWAQYEQD | TP | 1 | Homo sapiens (Human) | |
| Q96I34 PPP1R16A PP16A_HUMAN |
30 | 37 | KHAQKRRAQQVKMWAQAEKE | TP | 1 | Homo sapiens (Human) | |
| O60237 PPP1R12B MYPT2_HUMAN |
19 | 26 | ESARMRRAEQLRRWRGSLTE | TP | 1 | Homo sapiens (Human) | |
| O14974 PPP1R12A MYPT1_HUMAN |
10 | 17 | ADAKQKRNEQLKRWIGSETD | TP | 1 | Homo sapiens (Human) | |
| Q9Y6X6 MYO16 MYO16_HUMAN |
24 | 31 | RLVKRMRCEQIKAYYEREKA | TP | 1 | Homo sapiens (Human) |
Please cite:
ELM-the Eukaryotic Linear Motif resource-2024 update.
(PMID:37962385)
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement