| Accession: | |
|---|---|
| Functional site class: | PAM2 motif |
| Functional site description: | The PABP-interacting motif 2 (PAM2) mediates binding of proteins to the MLLE/PABC peptide-binding domain found in poly(A)-binding proteins and HYD E3 ubiquitin ligases. There are two variants of the PAM2 motif that bind to overlapping but distinct sites on the MLLE domain, adopt a different orientation at their termini and consequently have different recognition elements in these regions. While the core region around a critical phenylalanine is similar in the LIG_PAM2_1 and LIG_PAM2_2 variants and makes identical contacts with the MLLE domain, the N-terminal sequence in the LIG_PAM2_1 provides a hydrophobic residue that is essential for efficient binding and is lacking in LIG_PAM2_2 motifs, while the C-terminal part of the LIG_PAM2_2 variant makes a turn and follows an alternative path on the MLLE surface that depends on an aromatic residue that is absent from LIG_PAM2_1 motifs. |
| ELMs with same func. site: | LIG_PAM2_1 LIG_PAM2_2 |
| ELM Description: | Binding of the LIG_PAM2_2 mainly involves hydrophobic interactions, the most important of which are mediated by the residues at position 5 (corresponds to position 10 in LIG_PAM2_1) and 11 (lacking in LIG_PAM2_1). A first set of hydrophobic contacts is provided by the residues in positions 1 and 2, which are preferably tryptophan and proline, respectively. However, the motif in D. melanogaster GW182, which binds with a lower affinity to the MLLE domain, contains other hydrophobic residues in these positions, similar to other invertebrates (phenylalanine or leucine and proline or valine observed in position 1 and 2, respectively). The invariant phenylalanine in position 5 (corresponds to position 10 in LIG_PAM2_1) is the most important residue for binding, together with the tryptophan in position 11, which is lacking in the LIG_PAM2_1 variant. The beta-turn conformations adopted by the proline and glycine in positions 7 and 8 and the lysine and glycine residues in positions 12 and 13 are stabilized by intra- and intermolecular hydrogen bonds, which might explain the preference for these residues in these positions. The proline in position 7 provides an additional hydrophobic contact with the MLLE domain. The invariant glutamate in position 4 forms a salt bridge with an MLLE lysine residue. Structural analysis (3KTP) indicates that the last motif residue also provides a hydrophobic contact, and indeed in vertebrate sequences this position is occupied by a limited set of hydrophobic residues. However, in invertebrates the preference is less strict, and since the motif definition is already quit strict, this position is currently defined as a wildcard. The motif seems to be quite specific and contains several invariant residues. Although its binding partner is present in other eukaryotes, the GW182 protein family seems to be restricted to metazoans. Hence, this variant of the PAM2 motif seems to be specific for animals, and other eukaryotic groups might use distinct variants. |
| Pattern: | ((WPP)|([FL][PV][APQ]))EF.PG.PWKG. |
| Pattern Probability: | 9.773e-14 |
| Present in taxon: | Metazoa |
| Interaction Domain: |
PABP (PF00658)
Poly-adenylate binding protein, unique domain
(Stochiometry: 1 : 1)
|
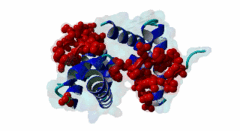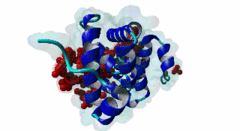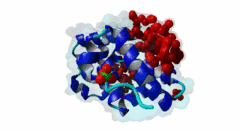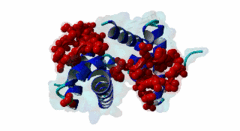
The PABP-interacting motif 2 (PAM2) directly binds to the MLLE/PABC domain that is found in poly(A)-binding proteins (PABP) like PABPC1 and in members of the HECT domain-containing Hyperplastic Discs (HYD) protein family of E3 ubiquitin ligases (Albrecht,2004, Lim,2006). This domain consists of a conserved bundle of five alpha-helices, of which the N-terminal helix is lacking in some proteins. The interaction with PAM2 sequences involves the most conserved helices 2, 3 and 5, with the exception of yeast, where only helices 2 and 3 mediate binding. PABPC1 binds to the 3’-poly(A) tail of mRNA molecules via four RNA recognition motifs (RRMs) and recruits different regulatory proteins, which modulate translational activity and mRNA stability, by binding to their PAM2 motifs via its C-terminal MLLE domain (Siddiqui,2007). LIG_PAM2_1 motif-containing PABPC1-binding proteins include Paip1 and Paip2, which stimulate or repress translation by stabilizing or destabilizing, respectively, the closed loop structure of mRNA that is formed by the interaction between PABP and the translation initiation factor eIF4G. Other examples are the eukaryotic release factor eRF3, which contains two overlapping LIG_PAM2_1 motifs, and Tob1 and Pan3, which recruit the deadenylase complexes Caf1-Ccr4 and Pan2-Pan3, respectively, to the mRNA poly(A) tails. These complexes are involved in translation-dependent, eRF3-mediated mRNA decay and translation termination. Tob1 and its family member Tob2 contain two distinct LIG_PAM2_1 motifs, and evidence indicates that in both cases the C-terminal motif is the main interaction site for binding to PABP. PABPC1 binding partners containing the LIG_PAM2_2 motif variant are so far restricted to animal-specific GW182 family proteins, which are part of the RNA-induced silencing complex (RISC) and are essential for microRNA (miRNA)-mediated gene silencing (Jinek,2010). Silencing is achieved by inhibiting translation and promoting degradation of mRNA, and depends on binding of GW182 proteins in the miRNA-loaded RISC to PABPC1. Additional, possibly indirect, interactions also occur between the RRMs of PABPC1 and the C-terminal region of GW182 family members. While in human GW182 proteins PABPC1 binding is predominantly mediated by their PAM2 sequences, in D. melanogaster GW182 the motif, which binds with lower affinity, also contributes to but is dispensable for this interaction in immunoprecipitation experiments (Huntzinger,2010). The tumor suppressor protein HYD (EDD/UBR5) belongs to the family of HECT domain-containing E3 ubiquitin ligases, which target specific proteins for proteasome-dependent degradation. Evidence suggests that HYD is involved in proliferation and DNA damage signaling. Little is known about the function of the MLLE-PAM2 interaction in HYD E3 ligases, but it has been suggested that PAM2-containing proteins may be targeted for ubiquitination by HYD through binding to its MLLE domain, which is positioned directly adjacent to the catalytic HECT domain. So far, all known PAM2-mediated interactions with HYD involve instances of the LIG_PAM2_1 variant. The structural features of PAM2 binding to the MLLE domain of PABP or HYD are very similar, and similar specificity and binding affinity has been demonstrated for various peptides (Lim,2006). |
(click table headers for sorting; Notes column: =Number of Switches, =Number of Interactions)
| Acc., Gene-, Name | Start | End | Subsequence | Logic | #Ev. | Organism | Notes |
|---|---|---|---|---|---|---|---|
| Q9HCJ0 TNRC6C TNR6C_HUMAN |
1385 | 1398 | SINWPPEFHPGVPWKGLQNI | TP | 15 | Homo sapiens (Human) | |
| Q8SY33 gw GAWKY_DROME |
957 | 970 | YTDLVQEFEPGKPWKGSQIK | TP | 4 | Drosophila melanogaster (Fruit fly) | |
| Q9UPQ9 TNRC6B TNR6B_HUMAN |
1476 | 1489 | NASWPPEFQPGVPWKGIQNI | TP | 4 | Homo sapiens (Human) | |
| Q8NDV7 TNRC6A TNR6A_HUMAN |
1608 | 1621 | SVNWPPEFRPGEPWKGYPNI | TP | 2 | Homo sapiens (Human) |
Please cite:
ELM-the Eukaryotic Linear Motif resource-2024 update.
(PMID:37962385)
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement
ELM data can be downloaded & distributed for non-commercial use according to the ELM Software License Agreement